REACH-Ab
Antibody design, guided from the first edge.
A dynamic interface field that lets epitope-paratope contacts emerge before the paratope geometry is already fixed. Designed for sequence, structure, and binding interface quality to improve together.
Highlights
First, the essentials.
CDR-H3 AAR on RAbD.
LDDT for generated CDR-H3 structure.
DockQ for single CDR-H3 generation.
Fifty-seven GIFs reveal how edges emerge, compete, persist, and vanish.
Broad candidate field
Possible contacts stay visible before current geometry can reliably select them.
Dynamic edge memory
Weak evidence can accumulate instead of disappearing after one uncertain step.
Interface feedback
Selected relations feed back into local sequence and structure generation.
Preparing structure view...
Scientific Problem
Epitope guidance lag: when the right information arrives after the generative decision has begun
The central claim of REACH-Ab is not that previous methods ignore the antigen. The sharper problem is that antigen information often reaches CDR generation through edges induced by a provisional paratope geometry. This makes epitope guidance posterior to the current CDR hypothesis, even though the epitope should help organize that hypothesis from the beginning.
Core contradiction
Paratope formation needs epitope contacts for guidance, but epitope contacts are often constructed from a paratope geometry that has not yet been reliably formed.
Design is interface organization, not only loop plausibility
Without a target epitope, many CDR sequences and conformations can look locally reasonable. With a target epitope, the model must generate a paratope that realizes a binding-compatible cross-interface contact pattern. A plausible CDR that misses the antigen-facing organization is still a failure for epitope-conditioned design.
Nodes are persistent, while interface edges are often derived
Many geometric generators maintain residues, atoms, frames, or coordinates as the main evolving objects. Cross-interface edges are then rebuilt from current distances, kNN neighborhoods, pairwise scores, or attention-like links. These edges can pass antigen information, but they remain consequences of the current node hypothesis rather than independent contact hypotheses.
Early geometry becomes the gatekeeper of epitope information
When the CDR is still uncertain, the model most needs epitope contacts to constrain generation. Yet if contact edges are induced only from provisional geometry, then early local shape decides which epitope residues are visible, active, and able to influence the next update. Useful future contacts can be delayed or excluded before the paratope has had a fair chance to form.
Generation becomes path-dependent
Incorrect but geometrically close contacts can be reinforced, while correct but not-yet-close contacts may be ignored. Later refinement may still improve local structure, but the search has already been narrowed by early provisional geometry. This is why final loop plausibility can coexist with weak contact recovery, high interface RMSD, or low DockQ.
The problem must be observed along the trajectory
Because epitope guidance lag is a timing problem, final metrics alone are not enough. The process should reveal whether correct contacts are visible early, whether they become active edges early, whether they emerge before the final geometry is settled, and whether wrong active edges persist across refinement.
Edges need anticipation, persistence, and feedback
A useful interface edge should be able to participate before the paratope is stable, keep memory across refinement steps, and carry relation content back into node updates. Conventional dynamic graphs may change over time, but if every change is still induced by current node geometry, the edges remain reactive rather than generative.
Node-centric timing
Generate or update CDR nodes → induce interface edges from provisional geometry → pass antigen information.
REACH-Ab timing
Maintain candidate interface edges early → update edge states with memory and compatibility → feed selected edge states back into generation.
Result
Quantitative evidence from the main paper and supplementary experiments
The tables below collect the experimental result tables from the manuscript and supplementary material. Bold cells mark the best result in the corresponding comparison, while underlined cells mark the second-best result when reported in the paper.
Main Paper
Task-level performance
CDR-H3 generation on RAbD
Generation quality and interface-sensitive evaluation for epitope-conditioned CDR-H3 design.
| Model | Generation | Interface | ||||
|---|---|---|---|---|---|---|
| AAR ↑ | TM-score ↑ | LDDT ↑ | CAAR ↑ | RMSD ↓ | DockQ ↑ | |
| RosettaAb | 32.14% | 0.9416 | 0.8002 | 16.79% | 17.7735 | 0.197 |
| HERN | 31.59% | -- | -- | 20.09% | 11.1451 | 0.057 |
| DiffAb | 26.46% | 0.6087 | 0.5665 | 20.07% | 19.1944 | 0.119 |
| MEAN | 33.50% | 0.7855 | 0.5616 | 22.11% | 19.4989 | 0.132 |
| ADesigner | 34.40% | 0.6321 | 0.5154 | 21.90% | 10.7468 | 0.225 |
| dyMEAN | 35.13% | 0.8579 | 0.6668 | 20.24% | 11.5814 | 0.325 |
| dyAb | 35.91% | 0.8593 | 0.6557 | 22.45% | 10.8222 | 0.330 |
| REACH-Ab | 40.76% | 0.9787 | 0.8750 | 25.31% | 7.7024 | 0.431 |
Interpretation
CDR-H3 is the most variable antigen-facing loop, so this benchmark tests whether a model can improve local loop generation while also forming the correct antigen-facing interface. Several baselines recover plausible H3 structures, but their CAAR and DockQ remain limited, showing that a reasonable loop shape can still miss the intended contact pattern.
REACH-Ab leads both the generation metrics and the interface-sensitive metrics. This is the key point: the edge field does not simply trade local CDR quality for docking quality. Instead, active interface hypotheses constrain H3 refinement inside an antigen-aware contact space, which improves sequence, structure, and interface organization at the same time.
Antibody-antigen complex structure prediction
Pipeline-based methods are separated from end-to-end deep learning models.
| Method | TM-score ↑ | LDDT ↑ | RMSD ↓ | DockQ ↑ |
|---|---|---|---|---|
| IgFold->Hdock | 0.9706 | 0.5398 | 17.65 | 0.228 |
| IgFold->HERN | 0.9719 | 0.5488 | 16.16 | 0.222 |
| dyMEAN | 0.8919 | 0.6366 | 10.97 | 0.392 |
| dyAb | 0.9079 | 0.6610 | 10.91 | 0.412 |
| REACH-Ab | 0.9698 | 0.8593 | 9.18 | 0.475 |
Interpretation
Complex prediction separates antibody fold recovery from antibody-antigen assembly. The IgFold-based pipelines maintain strong TM-score, yet their LDDT, RMSD, and DockQ are much weaker, indicating that a correct antibody fold alone does not guarantee a correct binding interface.
REACH-Ab achieves the strongest local and interface-sensitive scores among the compared methods. This supports the timing argument behind the method: candidate interface edges are maintained during refinement, selected edge states are fed back into node updates, and unstable contact hypotheses can be suppressed before they distort the final complex.
Affinity optimization under mutation budgets
Mutation budgets are 1, 2, 4, 8, and all mutable positions.
| Metric | Model | 1 | 2 | 4 | 8 | All |
|---|---|---|---|---|---|---|
| ΔΔG ↓ | MEAN | 0.00 | 0.00 | -0.16 | -3.29 | -4.01 |
| ADesigner | -0.89 | -1.46 | -1.9558 | -2.47 | -2.64 | |
| dyMEAN | -3.01 | -3.59 | -3.7599 | -4.05 | -4.95 | |
| dyAb | -3.15 | -4.74 | -4.819 | -5.19 | -5.76 | |
| REACH-Ab | -3.25 | -3.43 | -3.63 | -3.70 | -3.81 | |
| ΔL | MEAN | 0.00 | 0.00 | 0.75 | 7.91 | 10.89 |
| ADesigner | 0.78 | 1.18 | 3.54 | 7.17 | 10.24 | |
| dyMEAN | 0.97 | 1.53 | 3.45 | 6.55 | 9.02 | |
| dyAb | 0.91 | 1.75 | 3.45 | 7.79 | 8.81 | |
| REACH-Ab | 1.00 | 1.51 | 2.67 | 5.49 | 8.80 |
Interpretation
Affinity optimization is a different regime from generation or complex prediction. It rewards useful deviation from an existing complex, while still requiring the mutated antibody to preserve a credible binding interface. For this reason, both predicted affinity change and mutation aggressiveness matter.
REACH-Ab is competitive under the one-mutation budget, but its affinity gain saturates as the budget increases and its mutation distance remains comparatively conservative. This exposes a useful boundary of the current mechanism: persistent edge states are strong at preserving coherent interface geometry, but affinity search may require controlled exploration away from the original contact basin.
Mechanism inset from edge-field-guided generation
Process-level metrics compare REACH-Ab with a geometry-induced kNN baseline.
| Metric | Baseline | REACH-Ab | Gain |
|---|---|---|---|
| CVG ↑ | 44.5% | 100.0% | +55.5 pp |
| CAG ↑ | 44.5% | 60.6% | +16.0 pp |
| CEL ↑ | 71.6% | 81.0% | +9.4 pp |
| FPRR ↓ | 67.1% | 47.5% | +19.6 pp |
Interpretation
Final structure scores show whether the output is good, but they do not reveal when epitope guidance becomes available during generation. This inset therefore evaluates the process itself: whether contact evidence is visible early, becomes active early, emerges earlier across refinement, and avoids persistent false-positive reinforcement.
REACH-Ab improves contact visibility, contact activation, and cumulative contact emergence while also reducing false-positive persistence. This combination is important: the broad edge field is not merely adding more noisy edges. It keeps useful candidates available, selects a better active subset, and prevents unsupported contacts from dominating message passing.
Key ablation on epitope-conditioned CDR-H3 generation
Variants remove edge-field components while keeping parameter scale comparable where possible.
Generation metrics
| Variant | AAR ↑ | TM-score ↑ | LDDT ↑ |
|---|---|---|---|
| REACH-Ab | 40.76% | 0.9787 | 0.8750 |
| w/o edge field | 38.75% | 0.9731 | 0.8665 |
| w/o broad space | 38.36% | 0.9271 | 0.8604 |
| w/o half-edge | 39.02% | 0.9363 | 0.8101 |
| w/o edge feedback | 38.52% | 0.9565 | 0.8105 |
Interface metrics
| Variant | CAAR ↑ | RMSD ↓ | DockQ ↑ |
|---|---|---|---|
| REACH-Ab | 25.31% | 7.7024 | 0.431 |
| w/o edge field | 23.37% | 8.3241 | 0.422 |
| w/o broad space | 24.16% | 8.9556 | 0.407 |
| w/o half-edge | 23.34% | 9.1534 | 0.398 |
| w/o edge feedback | 24.01% | 10.2957 | 0.326 |
Interpretation
The full model performs best across both generation and interface metrics, which indicates that the improvement comes from the edge-field mechanism as a system rather than from a single isolated score adjustment. Removing the full edge field mainly weakens interface organization, even when some structure metrics remain strong, so the lost capability is not just backbone plausibility but antigen-aware contact formation.
The component ablations clarify what each part contributes. Removing the broad candidate space weakens geometry-sensitive metrics, showing that early interface search cannot be reduced to current-distance neighborhoods. Removing half-edge probing lowers LDDT and DockQ, suggesting endpoint compatibility is needed for stable interface geometry. Removing edge-state feedback gives the sharpest DockQ and RMSD degradation, which means selected edges must carry compatibility, uncertainty, and memory into message passing rather than serving only as binary links.
Supplementary Material
Extended result tables
Seed sensitivity analysis
Mean and standard deviation across seeds for epitope-conditioned CDR-H3 generation.
| Model | Generation mean ± std | Interface mean ± std | ||||
|---|---|---|---|---|---|---|
| AAR ↑ | TM-score ↑ | LDDT ↑ | CAAR ↑ | RMSD ↓ | DockQ ↑ | |
| RosettaAb | 32.14% | 0.9416 | 0.8002 | 16.79% | 17.7735 | 0.1968 |
| HERN | 31.64% ± 0.62% | -- | -- | 18.21% ± 1.53% | 10.49 ± 0.34 | 0.051 ± 0.004 |
| DiffAb | 29.32% ± 2.33% | 0.6359 ± 0.0181 | 0.5692 ± 0.0057 | 18.71% ± 1.32% | 17.74 ± 0.92 | 0.137 ± 0.013 |
| MEAN | 35.22% ± 0.91% | 0.8035 ± 0.0140 | 0.5554 ± 0.0144 | 20.94% ± 0.73% | 18.28 ± 0.85 | 0.112 ± 0.011 |
| ADesigner | 34.23% ± 1.78% | 0.6508 ± 0.0101 | 0.5042 ± 0.0066 | 20.73% ± 1.75% | 10.11 ± 0.45 | 0.246 ± 0.012 |
| dyMEAN | 35.52% ± 0.39% | 0.8554 ± 0.0035 | 0.6618 ± 0.0055 | 20.05% ± 0.12% | 11.77 ± 0.16 | 0.317 ± 0.005 |
| dyAb | 36.22% ± 0.41% | 0.8836 ± 0.0240 | 0.6662 ± 0.0151 | 23.22% ± 0.56% | 10.21 ± 0.46 | 0.329 ± 0.010 |
| REACH-Ab | 39.67% ± 0.62% | 0.9724 ± 0.0079 | 0.8608 ± 0.0088 | 24.28% ± 0.79% | 7.990 ± 0.17 | 0.424 ± 0.008 |
Interpretation
This table separates average performance from seed sensitivity. A method that improves only under a favorable initialization would show large variance, especially on docking-oriented metrics where small H3 geometry changes can strongly affect interface quality.
REACH-Ab remains best on the mean generation metrics and on the interface metrics, with modest standard deviations. This stability is important for the mechanism claim: the improvement is not a lucky final correction, but a repeatable effect of constraining refinement through persistent interface hypotheses.
Six-CDR co-design
Per-CDR AAR and global structure/interface metrics when all CDRs are generated together.
| Metric | Region | dyMEAN | dyAb | REACH-Ab |
|---|---|---|---|---|
| AAR ↑ | H1 | 56.32% | 48.68% | 70.44% |
| H2 | 42.39% | 32.86% | 63.57% | |
| H3 | 33.51% | 23.50% | 39.69% | |
| L1 | 42.91% | 19.29% | 60.68% | |
| L2 | 45.86% | 41.69% | 77.27% | |
| L3 | 42.43% | 20.11% | 42.31% | |
| TM-score ↑ | -- | 0.8868 | 0.6667 | 0.9639 |
| LDDT ↑ | -- | 0.7849 | 0.6234 | 0.8341 |
| DockQ ↑ | -- | 0.4192 | 0.2416 | 0.4428 |
Interpretation
Six-CDR co-design is harder than single-H3 generation because the model must organize a full paratope instead of one dominant loop. As all CDRs become mutable, antigen-conditioned information has to be distributed across multiple loops that may compete for interface contacts.
REACH-Ab improves sequence recovery on five of the six CDRs and remains comparable on L3, while also achieving the best TM-score, LDDT, and DockQ. The result suggests that the dynamic edge field does not overfit to H3-specific behavior. It can maintain a shared set of epitope-paratope contact hypotheses and use selected edge states to coordinate the whole paratope during refinement.
Trajectory-level metrics for epitope guidance lag
Definitions used to quantify when native-like contact evidence becomes visible and active.
| Metric | Meaning | Reported quantity | Direction |
|---|---|---|---|
| CVG | Contact visibility | \(V_{\mathrm{R}}^0 - V_{\mathrm{K}}^0\) | Higher is better |
| CAG | Contact fraction | \(A_{\mathrm{R}}^0 - A_{\mathrm{K}}^0\) | Higher is better |
| CEL | Cumulative contact emergence lead | \(\frac{1}{T}\sum_t(E_{\mathrm{R}}^t - E_{\mathrm{K}}^t)\) | Higher is better |
| FPRR | False active-edge persistence rate | Method-specific rate | Lower is better |
How to read this table
These metrics turn epitope guidance lag into observable process-level quantities. CVG asks whether correct contact evidence is available in the candidate field at the beginning of refinement; CAG asks whether such evidence is already selected into the active graph and can transmit messages; CEL measures whether useful contacts appear earlier across the trajectory.
FPRR is the complementary failure check. A broad candidate field would be risky if it simply reinforced incorrect active edges, so lower FPRR indicates that REACH-Ab can keep candidate contacts visible without forcing noisy contacts to persist. Together, the four metrics evaluate timing, activation, and stability rather than only final output quality.
Edge-state intervention study
All variants keep selected topology fixed and modify only edge attributes used in message passing.
| Variant | AAR ↑ | CAAR ↑ | TM-score ↑ | LDDT ↑ | RMSD ↓ | DockQ ↑ |
|---|---|---|---|---|---|---|
| Full REACH-Ab | 40.76% | 25.31% | 0.9787 | 0.8750 | 7.7024 | 0.431 |
| Topology only | 36.24% | 20.75% | 0.9431 | 0.8617 | 15.1983 | 0.156 |
| Shuffled edge state | 38.54% | 22.77% | 0.9429 | 0.8696 | 10.6439 | 0.357 |
| Random edge state | 33.19% | 22.64% | 0.9560 | 0.8684 | 9.7629 | 0.380 |
Interpretation
This intervention isolates whether REACH-Ab benefits only from selecting a better active topology, or whether the learned edge states themselves carry generative information. All variants keep the selected active edge set fixed and modify only the edge attributes passed into message passing.
The topology-only setting produces the largest diagnostic drop, with DockQ falling from 0.431 to 0.156 and RMSD increasing from 7.70 to 15.20. Shuffled and random edge states also degrade performance, showing that useful feedback requires edge states that are learned and matched to the active contact hypothesis they represent. In other words, REACH-Ab is not only choosing which residues may interact; it is also passing dynamic relation content back into sequence and structure generation.
Mechanism Figures
How REACH-Ab turns epitope guidance into evolving interface relations
These figures summarize the scientific story behind the viewer: REACH-Ab first defines the antibody design task at the complex level, then maintains candidate epitope-paratope relations as dynamic states, and finally feeds selected relation states back into local message passing.
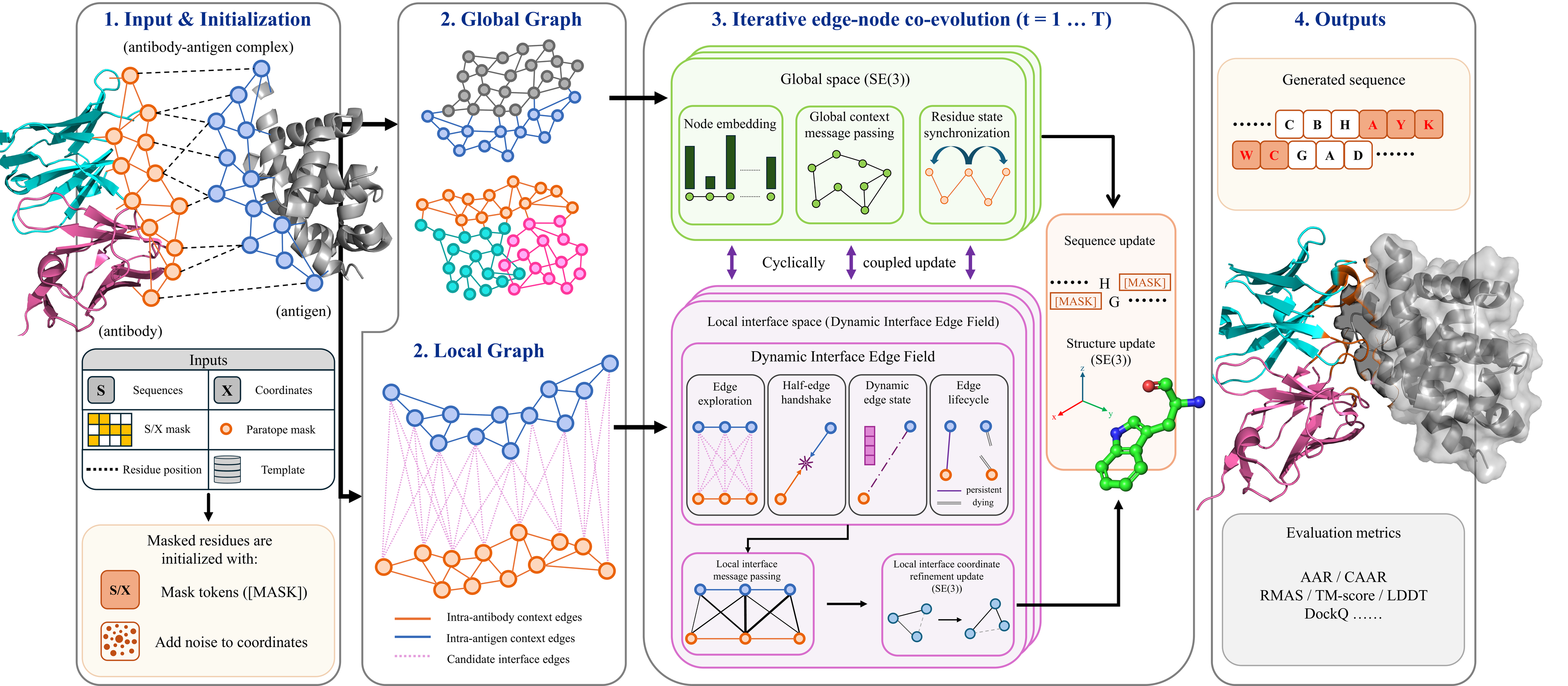
Framework Overview
From masked CDR residues to generated antibody sequence and complex structure
The framework initializes masked CDR residues from an antibody-antigen complex, builds global and local graphs, and iteratively couples SE(3) message passing with a local Dynamic Interface Edge Field. The key idea is that candidate interface edges are explored, updated, and selected before they guide local coordinate refinement and sequence generation.
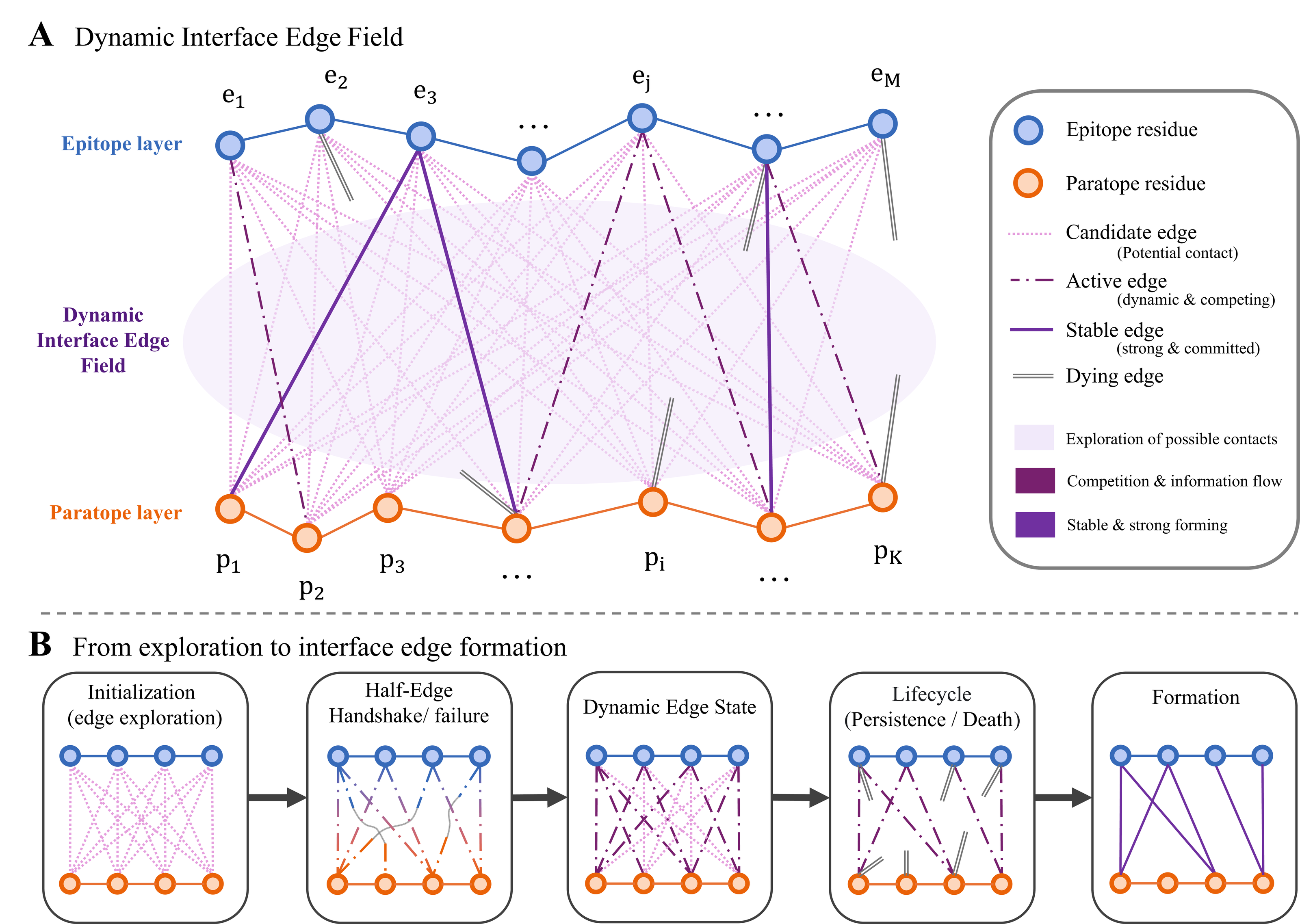
Core Concept
A dense relation field becomes a sparse active interface graph
REACH-Ab maintains many possible epitope-paratope contacts instead of committing only to current geometric neighbors. During refinement, half-edge evidence and lifecycle control let some relations stabilize, some compete, and some die out. This is the mechanism that reduces epitope guidance lag: antigen-facing contact hypotheses remain available before the paratope geometry is fully reliable.
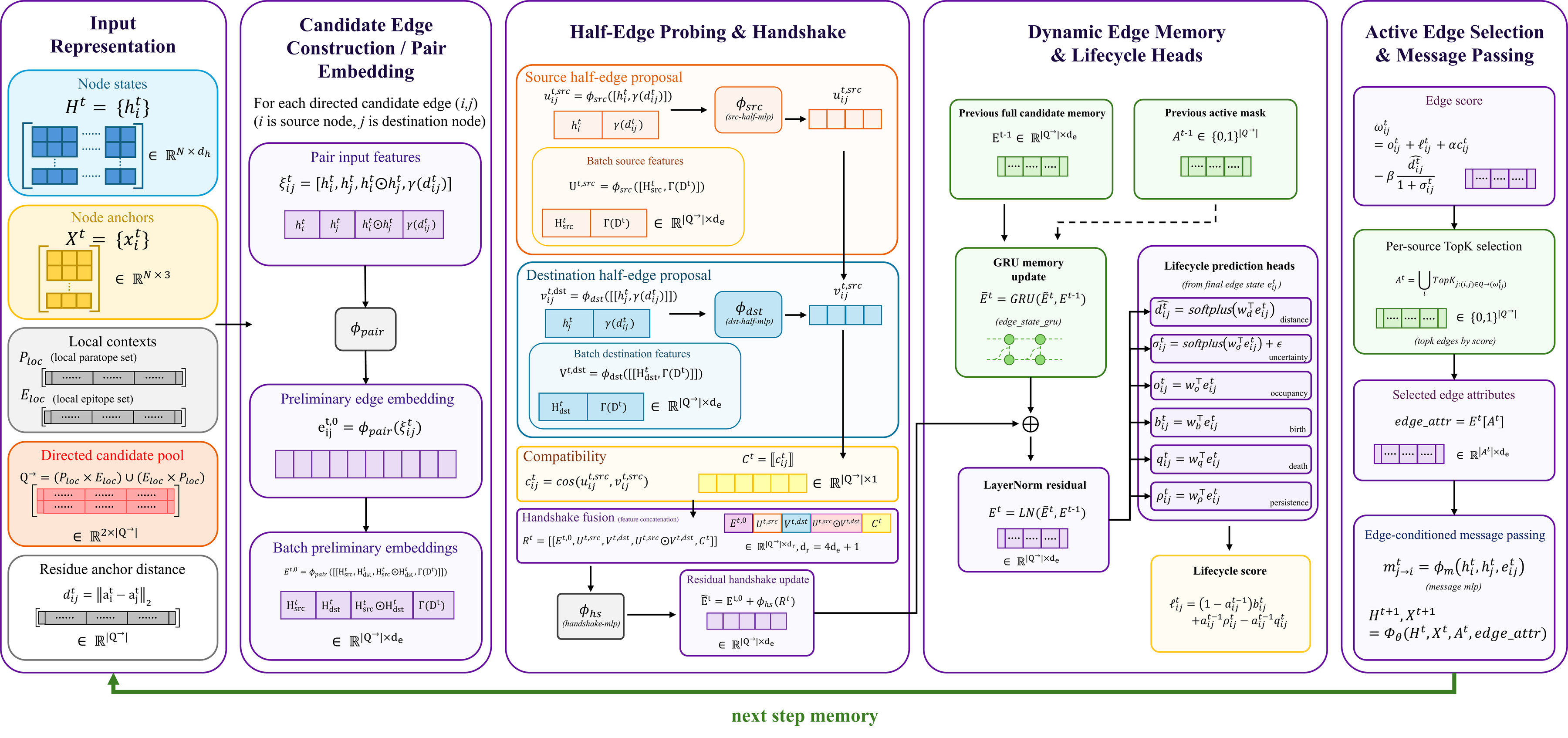
Implementation Flow
Candidate edges carry memory, compatibility, uncertainty, and lifecycle signals
At each refinement step, residue states and anchors construct a directed candidate edge pool. Pair-level features are updated through source-side and destination-side half-edge probing, then fused into persistent edge memory. Distance, occupancy, birth, death, persistence, and uncertainty heads produce an active edge score; selected edge states then become attributes in local message passing and are carried to the next refinement step.
Interactive Mechanism Playground
Feel the epitope guidance lag before reading about it
This toy generation sandbox turns the central mechanism into something visitors can manipulate. The left side rebuilds interface edges only from the current node geometry, while the right side maintains a broader edge field with memory and compatibility evidence before selecting active message-passing edges.
Early coordinates control geometry-kNN edges immediately; REACH-Ab keeps candidate contact hypotheses available while compatibility and memory evidence accumulates.
Edge-Field-Guided Generation
Watching contact hypotheses form during generation
The main-paper mechanism claim is temporal: REACH-Ab should expose useful epitope-paratope contact hypotheses before the final CDR geometry is settled. These 57 trajectories compare a geometry-kNN proxy with the REACH-Ab edge field for the same benchmark cases shown in the PDB viewer.
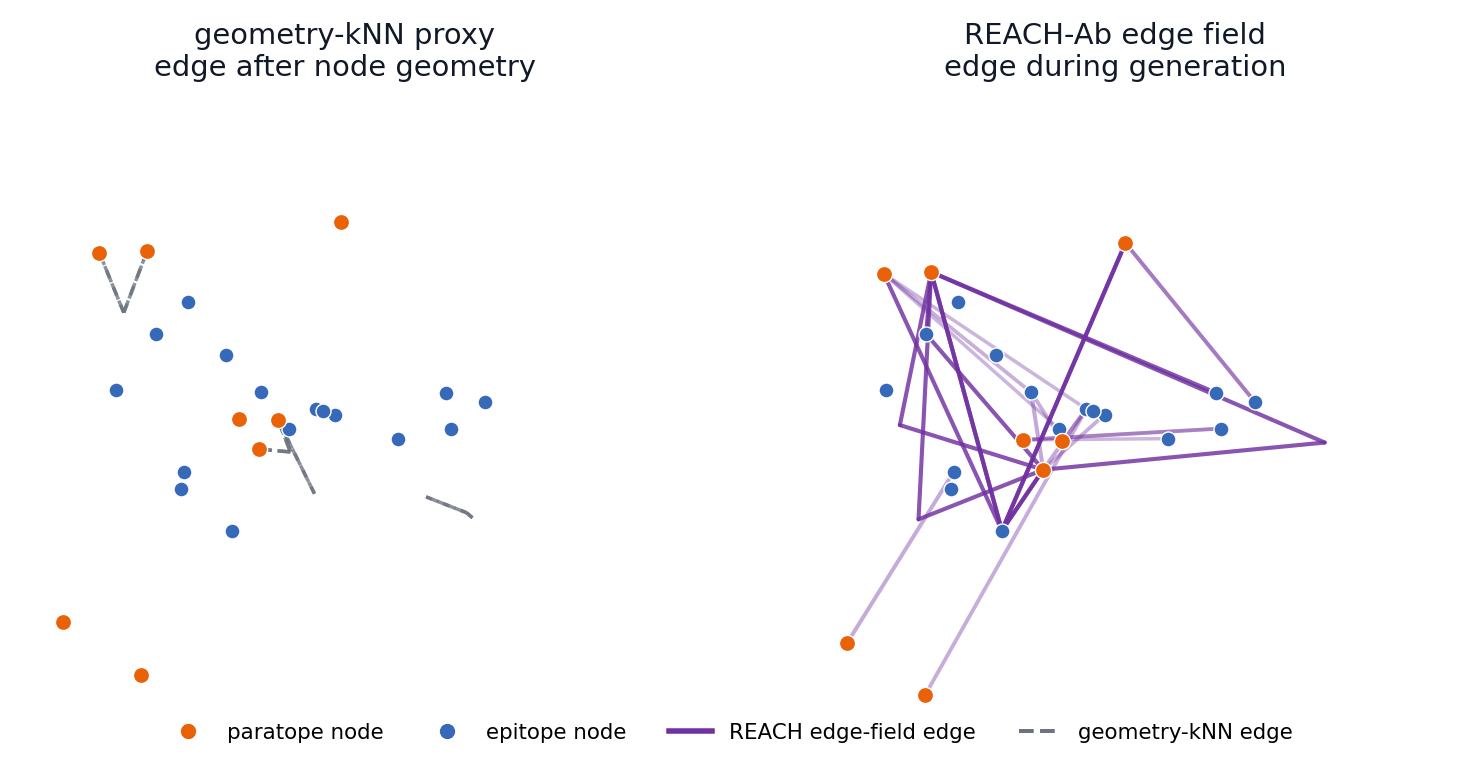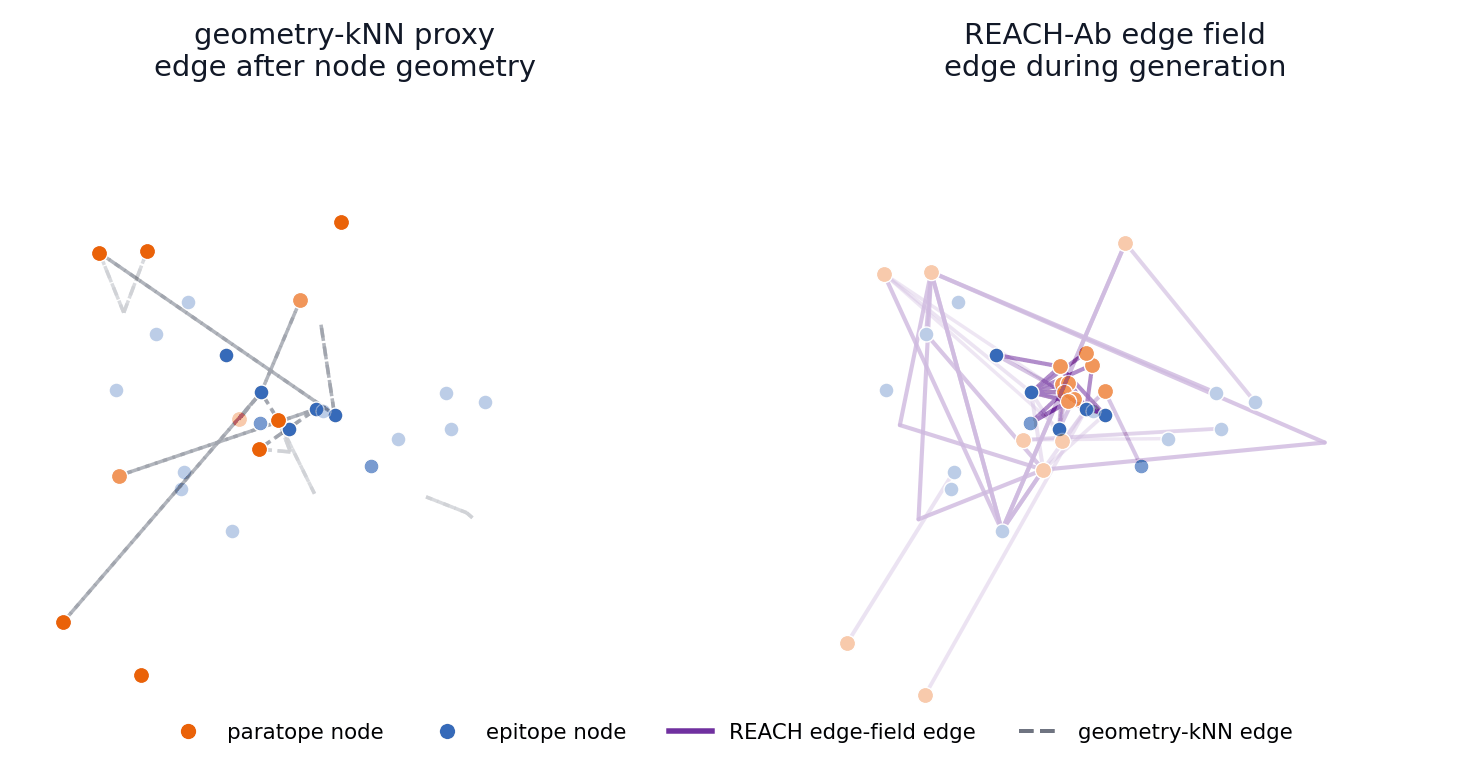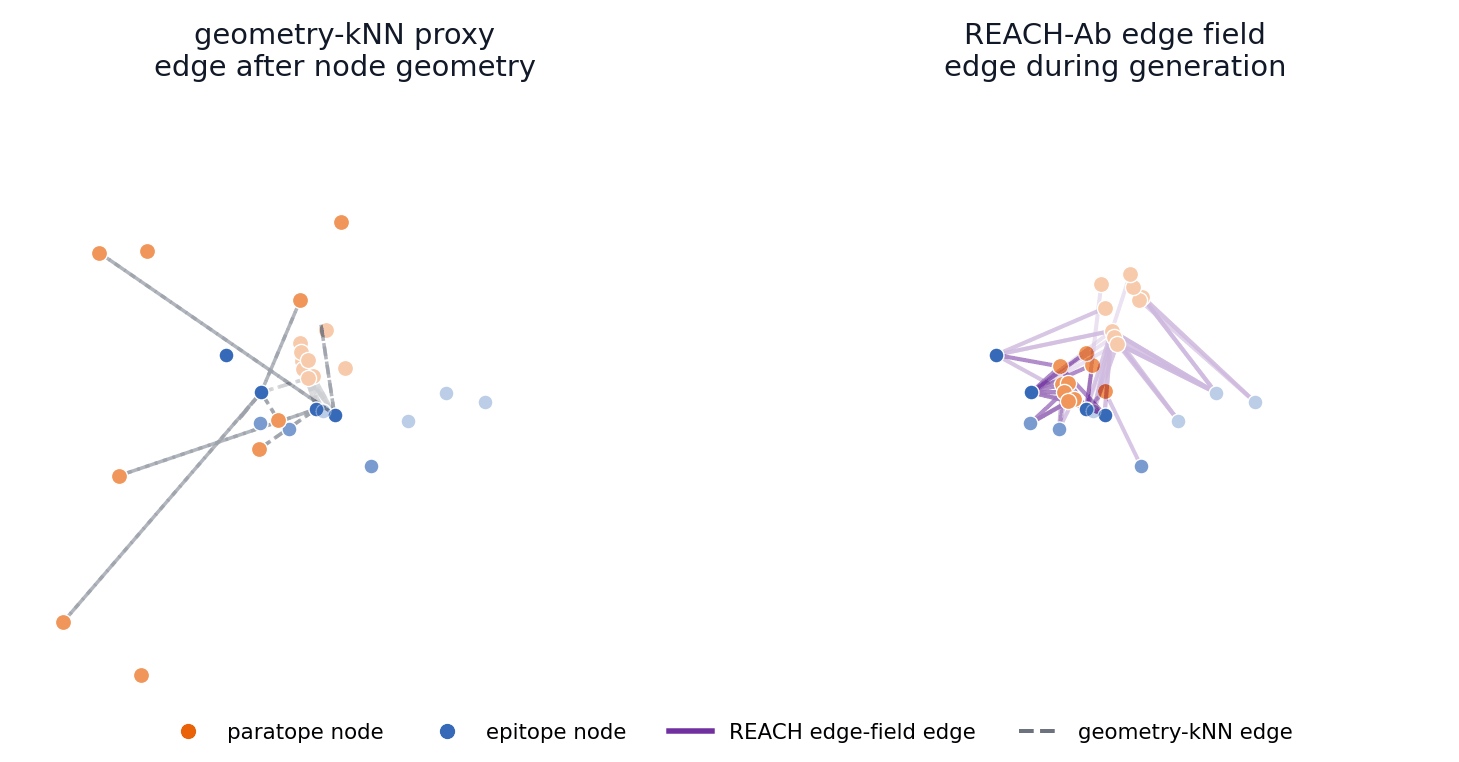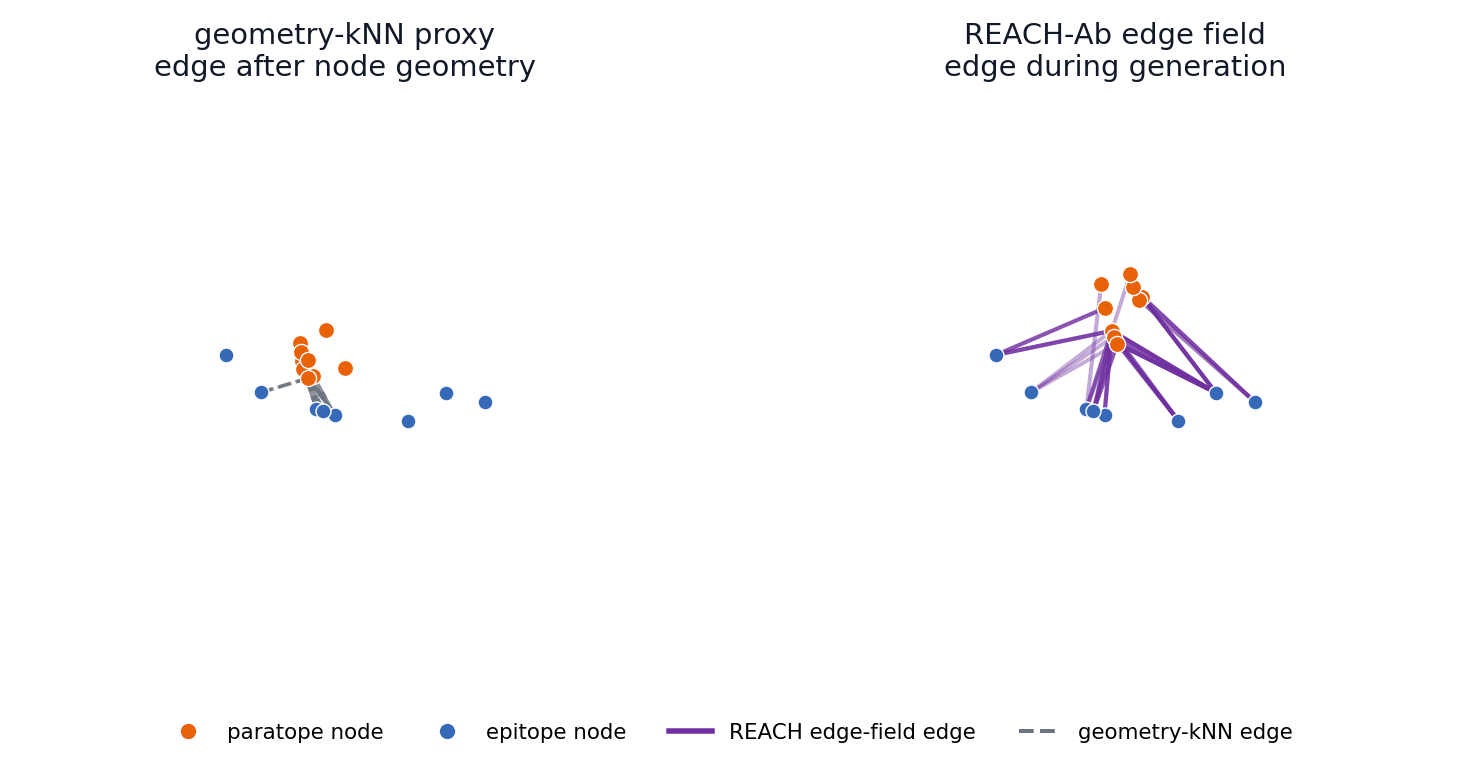
4FQJ edge-field trajectory
The left panel reconstructs edges after provisional node geometry appears. The right panel shows REACH-Ab maintaining and activating candidate edge states during refinement, so contact evidence can participate while the paratope is still being formed.
Paper to Code
Method equations linked to their implementation
This section merges the main-paper REACH-Ab method with the supplementary Model Architecture and Implementation Details and Theoretical Analysis. Each formula below is clickable: selecting it opens the concrete PyTorch implementation used by the current codebase.
Architecture Thread
REACH-Ab starts from a broad directed epitope-paratope candidate pool, encodes pair geometry and endpoint states, injects two-sided half-edge evidence, stores candidate-level memory, predicts lifecycle and uncertainty signals, then selects a sparse active interface graph for local message passing.
Theoretical Thread
The supplementary analysis explains why this matters: a broad candidate space preserves early contact accessibility, vector edge states are more expressive than scalar scores, half-edge probing generalizes one-sided scoring, and memory allows delayed activation from repeated weak evidence.